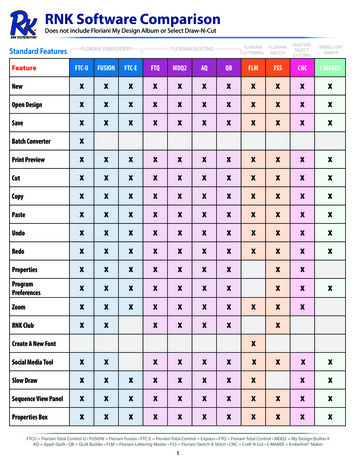
Transcription
SINTEZA RNK
Genska ekspresijaKako se jezik nukleotida prevodi u jezik proteina?
Genska ekspresijav Francis Crick (1957) postavio Centralnu DogmuMolekularne BiologijeØ Jednosmerni protokinformacijaØ Dvostepeni procesØ iRNK intermedijerOvaj proces se naziva genska ekspresija
Genska ekspresijaTranskripcija i translacija su sredstva pomoću kojihćelija čita ili eksprimira genetske instrukcije u svojimgenima.Mogu nastati brojne identične kopije RNK sa istoggena, a svaki RNK molekul može odrediti sintezumnogih identičnih molekula proteina.Ali svaki gen može biti transkribovan i preveden uprotein sa različitom efikasnošću, omogućavajući daćelija sintetiše velike količine jednih i male količinedrugih proteina.n
Geni mogu da se eksprimiraju sa različitom efikasnošćuGen A se više prepisuje od gena B. To obezbeđuje da protein A ućeliji ima utoliko veću koncentraciju u odnosu na protein B.
TranskripcijannNajveći broj gena u ćelijskoj DNK određuje sekvencuamino kiselina.RNK molekuli koji se kopiraju sa ovih gena senazivaju informaciona RNK (iRNK).Manji broj gena kao finalni proizvod daje samu RNK.
TranskripcijaTip RNKFunkcijaiRNKinformaciona RNK, kodira proteinerRNKribozomalna RNK, gradi osnovnu strukturu ribozomai katališe sintezu proteinatRNKtransportna RNK, uloga u sintezi proteina, adapterizmeđu iRNK i aminokiselinesnRNKmala jedarna RNK (small nuclear RNA), učestvuje ubrojnim procesima u jedru uključujući obraduisecanjem pre-iRNKsnoRNKmala nukleolarna RNK (small nucleolar RNA),učestvuje u obradi i hemijskoj modifikaciji rRNKDruge nekodirajuće Učestvuju u različitim ćelijskim procesima,uključujući sintezu telomera, inaktivaciju XRNKhromozoma i transport proteina u ER
Transkripcijav Pojedini regioni DNK se kopiraju u iRNKØiRNK je jednolančani molekulØ Jedan lanac DNK se čita: matrični lanacØ Drugi lanac liči na iRNK: ne-matrični lanacNe-matrični lanacDNKMatrični lanaciRNK
Transkripcijav Pojedini regioni DNK služe kao matrica za sintezuiRNKØMatrični lanac može da bude “gornji“ za jedan gen a“donji“ za drugi geniRNKNe-matrični lanacMatrični lanacDNKMatrični lanacNe-matrični lanaciRNKGen AGen BGen C
Transkripcijav Zašto se informacija sa DNK kopira na iRNK?Ø Štiti se “glavna kopija“ informacijeØ Proizvodi se iRNK samo za specifične geneØ Proizvodi se iRNK samo kada je potrebnaØ Nestabilnost iRNK omogućava regulacijuNe - matrični lanacDNKMatrični lanaciRNK
TranskripcijanEnzim koji vrši transkripciju jeDNK zavisna RNK polimeraza (RNKpolimeraza).nØRNK polimeraza katališe stvaranjefosfodiestarske veze koja povezujenukleotide u linearni lanac.RNK polimeraza eukariota:ooose sastoji od 12 subjedinica (uprokariota od 5).sadrži atome Zn koji činestrukturne komponente polimeraze isadrži atome Mg koji su prisutni uaktivnom mestu gde se odigravapolimerizacija.
Strukturna sličnost između bakterijske RNK poly u eukariotske RNK poly IISlični regioni su označeni zelenom bojomDodatni regioni su siviPlavo su atomi ZnCrveno su joni Mg-deo aktivnog centra gde se dešavapolimerizacijaZa razmotavanje DNK se ne troši energija-RNK poly i DNKkonformacione promene koje su energetski povoljne
TranskripcijaMatrični lanacDNKØ Sinteza novog lanca RNK se odvija u5‘ 3‘ pravcu.Ø Supstrati za RNK polimerazu sunukleozid trifosfati(ATP, CTP, UTP i GTP)Ø Hidroliza visokoenergetskih vezaobezbeđuje energiju neophodnu zaodvijanje ove reakcije.Ne-matrični lanac DNKØ Nukleozid 5‘ trifosfati se ređajukomplementarno bazama na jednomlancu DNK.SintezaNova RNK
Jedina razlika između replikacije DNK i transkripcije jestešto za transkripciju NIJE POTREBNA RNK POČETNICA.Oba procesa se odvijaju u 5‘ 3‘ pravcu, zahtevaju prisustvoaktiviranih nukleotida uz jone Mg i Mn koji ih činefunkcinonalnim
TranskripcijaTranskripcija se odvija u tri faze:Øinicijacija (započinjanje) lancaØ elongacija (produžavanje) lancaØ terminacija (završetak) lanca.Osnovni mehanizmi elongacije lanca u eukariota i prokariota su isti,dok se inicijacija i terminacija bitno razlikuju.
Transkripcija u prokariotav Inicijacija Promotor je sekvenca DNK koja predstavlja specifično mesto početkatranskripcije, u blizini 5’ kraja matričnog lanca gena.Sadrži oko 40 parova baza. Konsenzus sekvence - specifične sekvence baza u okviru promotora koje suvrlo slične kod različitih organizamaKonsenzus sekvenca 5’-TATAAT – 3’ se naziva Pribnow-ljeva struktura.KonsenzusØ 1 prva baza koja se prepisujeØ -10 i –35 sekvence definišu promotorPrimarni transkript
Transkripcija u prokariotav InicijacijaNajčešćeVećina faktora koji regulišu transkripciju gena vezuju se u blizinipromotora ili na samom promotoru i na taj načinutiču na inicijaciju transkripcije.
Transkripcija u prokariotav Inicijacija Transkripcija je najdetaljnije proučena kod E. coligde sva tri tipa RNK prepisuje samo jedna vrstaRNK-polimeraze.
Transkripcija u prokariotav RNK polimerazaØ Sastoji se iz pet subjedinica (polipeptidnih lanaca):α2, β , β, σ, koji čine holoenzim.Ø Tetramer α2, β , β naziva se kor-enzim.Ø Relativna molekulska masa celog enzima je oko 500.000daltona.“ kor“DNK
Transkripcija u prokariotav RNK polimerazaØ Sigma subjedinica RNK polimeraze se vezuje za -10/-35Ø Kor-enzim RNK polimeraze (α2β'β) vezuje sigmasubjedinicu (σ).“ kor“DNK
Transkripcija u prokariotav InicijacijaØ Rifampicin se vezuje za β-subjedinicu, kada se RNK polimerazanalazi u obliku holoenzima.Ø Vezivanjem za β-subjedinicu holoenzima rifampicin specifičnoinhibira inicijaciju transkripcije RNK.Ø Rifampicin nema uticaja na nuklearnu RNK eukariota.RNK polimeraza“kor-enzim"Promotorσ faktorRegion terminacije
Transkripcija u prokariotav ElongacijaØ RNK polimeraza otvara transkripcioni mehurićØ Započinje transkripcija matričnog lanca (bez početnice)Ø iRNK se sintetiše u 5' 3‘ smeruØ Oslobađa se sigma subjedinica (pošto se u molekul RNKveže oko 10 NT)ØUloga sigma subjedinice- prepoznavanje i vezivanje RNK poly zakorektno mesto inicijacije sinteze RNK lancaNe-matrični lanacNovosintetisana iRNKOslobođeni σ faktorMatrični lanacPravac transkripcije
Transkripcija u prokariotav ElongacijaØ Samo kratki delovi iRNK su preko baza spojeni sa DNKØ Iza RNK polimeraze se ponovo formira heliksTranskripcioni mehurićDNK se ponovo uvijaPromotorski regioniRNKSmer kretanjaRNK polimerazeRegionterminacije
Transkripcija u prokariotav ElongacijaØ Druga polimeraza može inicirati transkripciju čim seoslobodi promotorPromotorTranskripcija
Transkripcija prokariotav ElongacijaØ Druga polimeraza može inicirati transkripciju čim seoslobodi promotorNovosintetisana RNK sa proteinimaMolekuli RNKpolimerazeTerminacioni krajgenaInicijacioni krajgenaPravac sinteze
Transkripcija u eukariotav Klase RNK polimerazenKlasa enzimaRNKpolimeraza IOsetljivostprema ljiva45S RNKNukleolusRNKpolimeraza IIOsetljiva naniskukoncentracijuhnRNK (iRNK) NukleoplazmaRNKpolimeraza IIIOsetljiva navisokukoncentracijutRNK, 5SNukleoplazmaRNK, 7S RNKSpecifičan inhibitor eukariotske nukleoplazmatske RNK polimeraze II
RNK polimeraza IRNK polimeraza IIRNK polimeraza III
Transkripcija u eukariotav RNK polimerazaØRNK polimeraza eukariota, za razliku od RNK polimeraze prokariota,ne može da inicira transkripciju in vitro, već su potrebni dodatniproteini koji se nazivaju bazalni (osnovni) transkripcioni faktori.ØInicijacija transkripcije u eukariota mora rešiti problem pakovanjaDNK u nukleozome.Ø Bazalni transkripcioni faktori:1. pomažu postavljanje RNK polimeraze na promotor2. doprinose razdvajanju dva lanca DNK3. oslobađaju RNK polimerazu sa promotora u elongacionom modu,kada transkripcija počne.
Transkripcija u eukariotav Interakcija bazalnih transkripcionih faktora sa promotoromRNKpolimeraza IIRNKpolimeraza IIØ TATA boks je konsenzus sekvenca u promotoru koju prepoznajefaktor TFIID (sastoji se od TATA vezujućeg proteina (TBP) iTBP vezujućeg faktora).Ø Nalazi se 25 nukleotida uzvodno od mesta početka transkripcije.
Transkripcija u eukariotav Interakcija bazalnih transkripcionih faktora sa promotoromØ Vezivanje TFIID uzrokuje veliku distorzijuØØØØTATA boksa -označava se aktivni promotor.TFIIB se vezuje za TBP ise stvara kompleksTBP-TFIIB.Zajedno sa transkripcionim faktorom TFIIF,TFIIB služi kao most za vezivanje RNK polimeraze IITFIIH je sastavljen iz više subjedinica.- Prve dve subjedinice su helikaze kojeodvijaju DNK lanac na mestu početkatranskripcije- Sledeća subjedinica je protein-kinaza,fosforiliše ponavljajuće sekvenceC-terminalnog kraja RNK-polimeraze II.Fosforilacijom ovih sekvenci RNK-polimeraza IIse oslobađa iz kompleksa i nastavlja da se krećeduž DNK matrice i stvara RNK.
Kontrolni region tipičnog gena eukariotaPromotor je DNK sekvenca gde se vezuju bazalni faktori transkripcije ipolimeraza.Regulatorna sekvenca služi kao vezno mesto za regulatorni protein gena,čije prisustvo na DNK utiče na brzinu inicijacije transkripcije. Ovesekvence mogu da budu smeštene odmah uz promotor, daleko uzvodno od njegaili unutar introna ii nizvodno od samog gena.Smatra se da formiranje omče DNK omogućava regulatornim proteinima genada se vežu na bilo koju od pomenutih pozicija i da reaguju sa proteinima koji senalaze uz promotor.
Transkripcija u eukariotav InicijacijaØ Transkripcioni aktivatori su reglulatorni proteini gena koji pomažu upostavljanju RNK polimeraze na mesto početka transkripcije.Ø Medijatorni kompleks proteina omogućava da RNK polimeraza II komunicirasa bazalnim transkripcionim faktorima.Ø Kompleksi za remodelovanje hromatina i histon acetilaze takođe su čestopotrebni za inicijaciju RNK polimerazaiRNKGC boksCAT boksTATA boksRNKpolimeraza IITATA boksAdapterni molekul
Transkripcija u eukariotav InicijacijaPojačivačiGen specifične sekvence, dužine 1020 NT koji stimulišu transkripcijukod eukariota, bilo da su lokalizovanina početku ili na kraju gena alimoraju biti na istom lancu na komese nalazi i odgovarajući gen.Za razliku od promotora, pojačivačisu funkcionalni i na velikimudaljenostima od mesta početkatranskripcije (do 50 000bp).omogućeno postojanjem DNK petlje.Stimulacija transkripcije seostvaruje vezivanjem zatranskripcione faktore.
Transkripcija u eukariotav ElongacijaØ U toku elongacije za RNK polimerazu su vezani elongacioni faktoriproteini koji sprećavaju da RNK polimeraza disosuje pre nego štodostigne kraj gena.Ø Kompleksi za remodelovanje hromatina pomažu u kretanju RNKpolimeraze kroz nukleozome (i drugi elongacioni faktori verovatnoučestvuju u ovom procesu).Ø Problem stvaranja supernavoja u eukariota prevazilaze topoizomerazea u prokariota posebne topoizomeraze DNK giraze koje stvaraju negativnesupernavoje ispred mesta početka transkripcije.
Lokalna promena strukture hromatina koja jeuslovljena proteinima aktivatorima gena kod eukariotaAcetilacija histona i nukleazoma menja pakovanje hromatina činećiga pristupačnijim za vezivanje za druge proteine u ćeliji,uključujujući i proteine koje predstvaljaju inicijatore transkripije.
Dva specifična načina kojima acetilacija histonamože da stimuliše inicijaciju transkripcije(A) Neki proteini aktivatori mogu direktno da se vežu zaDNK koja je upakovana u nemodifikovani hromatin.Privlačenjem histon acetilaze (i kompleksa zaremodeliranje nukleazoma) ovi aktivatori olakšavajuvezivanje za DNK ostalih proteina aktivatora koji nemogu inače da se vežu za nemodifikovani hromatin. Oviproteini dalje mogu da uzrokuju modifikaciju hromatinaili deluju direktno kao transkripciona mašinerija.(B) Subjedinica opšteg transkripcionog faktora TFIIDsadrži dva kisela proteinska domena od kojih se svakisastoji od 120 AK, a nazivaju se bromodomeni. Svakibromodomen formira vezujući džep za jedan bočni lanacacetiliranog lizina (Ac);u TFIID ova dva džepa suodvojena sa 25 Å, što pretstavlja optimalno rastojanjeza prepoznavanje para acetiliranih lizina koji suodvojeni sa 6 ili 7 AK ostataka na N-terminalnom krajuhistona H4.Dodatno primeru acetilacije, ova subjedinica TFIIDtakođe prepoznaje N-terminalni kraj histona H4 kadaje acetilovan na poziciji 5 i 12 kao i potpuno acetiliranrep.On ne poseduje poseban afinitet prema neacetiliranomH4 repu i ima samo mali afinitet prema H4 repu koji jeacetiliran na samo jednom Liz.Različiti oblici acetilacije H4 su vezano zatranskripciono aktivne regione hromatina.
TranskripcijaØ Hromatin mora da bude de-kondenzovan§ Nukleozomi se ne odvijaju potpuno
Transkripcija–problem uvijanja dvostrukog heliksaTopoizomeraza I
Transkripcija – problem uvijanja dvostrukog heliksaTopoizomeraza II
Transkripcija u prokariotav TerminacijaPostoje dva načina terminacije:v terminacija zavisna samo od DNK-sekvence - ρ faktor nezavisnav terminacija pomoću proteina terminacije - ρ faktor zavisnaØ ρ faktor zavisna terminacija.- Pojedine sekvence imaju ulogu terminacionih sekvenci samo u prisustvu rofaktora.- Ro protein je heksamer molekulske mase 200.000.- Vezuje se za molekul RNK, i kreće prema molekulu RNK polimeraze, koja sezaustavlja na nekom terminacijskom mestu, istiskuje je sa 3' OH kraja RNK ioslobađa primarni transkript. Pri ovome se hidrolizuje jedan molekul ATP-a.
Transkripcija u prokariotav Terminacija-ρ faktor nezavisna terminacijaØ Terminaciona sekvenca u matričnom lancu DNK signalizirazavršetak sinteze molekula RNK.Ø Ispred mesta terminacije postoji sekvenca bogata citozinom iguaninom – palindrom sekvenca, koja sadrži međusobnokomplementarne regione, dolazi do savijanja RNK molekula iformiranja petlje u obliku ukosnice.Ø RNK polimeraza se zaustavlja i ponovo se formira DNK heliksTerminacioni regionTerminacioni signalUkosnica – terminacioni signaliRNKRNK polimerazaoslobođena saterminacione sekvence
Transkripcija u eukariotav TerminacijaTerminacija transkripcije kod eukariota obuhvata odvajanjenovonastalnog primarnog RNK transkripta uz adiciju Adeninskihnukleotida na 3' kraj transkripta.Ova poliadenilacija je potpuno nezavisna od matrice DNK sa kojese prepisuje primarni transkript RNK.Slično prokariotama, postoji tzv unutrašnja (Intrinsic) terminacija(Rho-nezavisna) pri čemu iRNK sadrži sekvence bogate C-Gparovima (7-20 bp), koje formiraju petlju. Ovu stabilnu petlju(trustruke H-veze između C-G) prati segment bogat uracilom pričemu su novoformirane veze između uracila i adenina slabe.Vezivanje RNK poly za petlju je sinhronizovano sa transkripcijompoli-uracil sekvence, tako da slabe Adenin-Uracil vezedestabilizuju RNA-DNA dupleks, i dovode do odvajanja RNK poly.
Obrada RNK
Zbir svih događaja od gena do proteina u bakterijama i eukariotskim ćelijamaKonačni nivo proteina u ćeliji zavisi od efikasnosti svakog koraka i od brzine razgradnje RNK i samogmolekula proteina.(A) U eukariotskim ćelijama, RNK koje nastaju transkripcijom sadrže u introne i egzone. Pre daljegtranslacije u protein, ovaj primarni transkript mora da se obradi, introni se uklanjaju enzimskikatalisanim reakcijama RNK “spicing-a” i tako dobijena RNK se iznosi iz jedra u citoplazmu.(B) U prokariotama, produkcija iRNK je mnogo jednostavnija. 5 kraj iRNK nastaje inicijacijomtranskripcije samom RNK poly, a 3 kraj nastaje terminacijom transkripcije.Kako prokariotske ćelije nemaju jedro, transkripcija i translacija se dešavaju u istom kompartmanu.Uglavnom translacija počinje po završetku transkripcije
Obrada RNK - prokarioteØ U prokariota većina molekula iRNK prepisanih sa matričnog lanca služi kaomatrica za sintezu JEDNOG proteina odmah po završetku transkripcije.Ø U prokariota iRNK je policistronska – sadrži prepis više gena koji noseinformaciju za nekoliko različitih genskih produkata.
Obrada RNKØ U eukariota primarni transkript tek nakon obrade postaje zrela iRNK.Ø Svi primarni transkripti u čijoj polimerizaciji učestvuje RNK polimeraza IInazivaju se heterogenom nuklearnom RNK (hnRNK ), jer su različite dužine.Ø Informaciona RNK eukariota nastaje obradom hnRNK.Ø Oko 3% hnRNK nizom modifikacijapostaje iRNK.Ø Obrada se obavlja primarno u jedru,ali i tokom transporta u citoplazmu.Ø Obrada iRNK odvija se u tri faze:§ modifikacija 5’ kraja§ modifikacija 3’ kraja§ isecanje i spajanje krajeva RNK.
Obrada RNKØ Obrada RNK je tesno povezana sa fazom elongacije transkripcije.Ø Ključni korak u preusmeravanju RNK polimeraze u elongacioni mod jeekstenzivna fosforilacija C-terminalnog kraja RNK polimeraze koji seskraćeno obeležava CTD (C-terminalni domen).Ø CTD se nalazi na najvećoj subjedinici i sastoji se od 52 puta ponovljenesekvence od 7 amino kiselina. Svaka sekvenca od 7 amino kiselina sadrži po 2 serina koja mogu bitifosforilisana. Kompletnom fosforilacijom polimeraze dodaje se ukupno 104 negativnonaelektrisanih fosfatnih grupa.
"RNK fabrika" RNK poly eukariotaNe obuhvata samo prepisivanje DNK u RNK većnosi sa sobom i proteine za obradu iRNK na svomrepu, koji se tada prenose na novosintetisamuRNK, u odgovarajućem trenutku.Postoji mnogo enzima koji su uključeni u obraduRNK i ne dolaze svi do RNK u sastavu RNK poly.Za RNK “splicing” na repu RNK poly se donosisamo nekoliko ključnih komponenti. Kada sejednom prenesu na RNK, oni predstavljaju centreza preostale komponente.Proteini za obradu RNK se prvo vezuju za RNKpoly rep pošto se on fosforiliše, kasno tokomprocesa inicijacije transkripcije. Kada RNKpoly IIzavrši transkripciju, odvaja se od DNK, uklanjajuse fosfatne grupe sa njenog repa i ona možeponovo da započne inicijaciju.Samo defosforilisana RNK poly može da iniciratranskripciju.
Obrada RNKØ Fosforilacija CTD RNK polimeraze:ð je odgovorna za disocijaciju RNK polimeraze II od drugih proteinaprisutnih na početnom mestu transkripcije i omogućava novoj grupiproteina, da se vežu za CTD polimeraze.- Ovi proteini učestvuju u elongaciji i obradi hnRNK.- Neki od njih prelaze sa RNK polimeraze na nascentni molekul RNKkoji počinje da se obrađuje neposredno nakon sinteze.Tako se, RNK polimeraza II u elongacionom modu može smatrati mašinerijomkoja i transkribuje DNK u RNK i obrađuje RNK neposredno nakon stvaranja.
Obrada RNK – modifikacija 5’ kraja5’ kraj novog RNK molekula se modifikuje:Ø neposredno nakon što RNK polimeraza II poveže oko 25 prvih nukleotidaØ dodavanjem "kape" koja se sastoji od modifikovanog guaninskog nukleotida.5’ kraj sadrži informacije koje se nikada ne prevode u proteine(vodeće sekvence) kao ni 3’ kraj (prateće sekvence)
5 -5 veza
Obrada RNK – modifikacija 5’ krajaU formiranju "kape" na 5’ kraju učestvuju 3enzima:1. Fosfataza – uklanja fosfat sa 5’ krajanascentne RNK2. Guanil transferaza – dodaje GMP sareverznom vezom (5’ – 5’ umesto 3’ – 5’)3. Metil transferaza – dodaje metil grupuguanozinu.Sva tri enzima se nalaze na CTD " repu “ RNKpolimeraze.Uloga 5’ kape:Ø razlikovanje iRNK od drugih tipova RNKØ vezivanje "kapa“ vezujućeg kompleksaproteina u jedru pomaže da se RNKadekvatno obradi i eksportuje iz jedra.Ø Učestvuje u translaciji iRNK u a
Obrada RNK – modifikacija 5’ krajaPostoje tri vrste “kapa” strukture u zavisnosti od toga da li su dvanukleotida koji su vezani za 7-metilguanilat metilisana.Ø “Kapa” 0 se javlja kada nema metilovanja iza 7-metilguanilata;Ø “kapa” 1 postoji kada je prva ribozna jedinica vezana za 7-metilguanilatmetilovana;Ø “kapa” 2 postoji kada su i prva i druga ribozna jedinica metilovane.Sve iRNK imaju “kapa” strukturu.
Obrada RNK – modifikacija 3’ krajaØ 3’ kraj iRNK je određen specifičnom sekvenecom u DNKØ Proteinið CstF (cleavage stimulating factor F) ið CPSF (cleavage and polyadenilation specificity factor)Putuju na “repu“ RNK polimeraze i prebacuju se na 3’ kraj čim se pojavispecifična sekvenca.- Za heksamer AAUAAA se vezuje CPSF.- Za GU bogatu sekvencu iza mesta cepanja se vezuje CstF.- CA sekvenca je treći faktor potreban za cepanje.
Obrada RNK – modifikacija 3’ krajaØ Posle vezivanja CPSF i CstF se vezuju se drugiproteini koji su uključeni u obradu 3’ kraja.Ø RNK se otcepljuje.Ø Poli A polimeraza dodaje jedan po jedan oko 200adeninskih nukleotida na 3’ kraj.- Prekursor je ATP.- Ne zahteva matricu.Ø Za poli A “rep“ se vezuje poli A- vezujući proteinkoji određuje definitivnu dužinu repa.Ø Poli A repð Pomaže u stabilizaciji iRNK.U citoplazmi, rep se polako skraćuje, i kadapotpuno nestane dolazi do brze degradacije iRNK.ð Takođe učestvuje u transportu iRNK iz jedra ucitoplazmu, i ima ulogu signala, na osnovu kogaribozomi prepoznaju odgovarajuću iRNK.
Obrada RNK – isecanje intronaØ Primarni prepis iRNK sadrži:ð kodirajuće sekvence - egzone, ið nekodirajuće sekvence – introne.Ø Introni su dugačke sekvencedeoksinukleotida (10 – 100,000 nukleotida)koje nemaju udela u genetskoj informacijikoja se prepisuje u redosled aminokiselinau polipeptidnom lancu.Ø Ove interventne sekvence postoje uvećini gena eukariota.Ø Introni variraju u veličini od 80-10 000nukleotida.
Obrada RNK – isecanje intronaØ Za razliku od sekvence egzona, tačnasekvenca introna je nevažna.Ø Jedino su sekvence koje učestvuju uuklanjanju introna, i koje se nalaze nakrajevima visoko konzervisane.Ø One su vrlo slične kod većine eukariota, isadržeð 5’ isecajuće/spajajuće mesto(donorsko mesto) ið 3’ isecajuće/spajajuće mesto(akceptorsko mesto).
Obrada RNK – isecanje intronaReakcije isecanja introna iz hnRNKØ Specifični adeninski nukleotid usekvenci introna napada 5’isecajuće/spajajuće mesto i sečešećerno-fosfatnu kičmu u ovoj tački.Ø 5’ kraj introna se kovalentno vezuje zaadeninski nukleotid.Ø Nastaje petlja (laso) u molekulu RNK.Ø Oslobođeni 3’ – OH kraj egzonareaguje sa početnom sekvencomsusednog egzona.Ø Dva egzona se povezuju u kontinuiranukodirajuću sekvencu.Ø Intron se oslobađa u obliku omče, irazgrađuje.
Obrada RNK – isecanje intronaIsecanje introna se odigrava u specifičnim česticama (engl. splicesome)Ø Isecanje introna se izvodi uz pomoć RNK molekula koji se nazivajumale nuklearne RNK – mnRNK (engl. small nuclear RNA - snRNA).Male nuklearne RNK – mnRNK:Ø Sadrže do 200 nukleotida.Ø Postoji pet vrsta: U1, U2, U4, U5 i U6.Ø Svaka mnRNK se povezuje sa 6-10 molekula proteina gradeći male nuklearneribonukleoproteine -mnRNP (small nuclear ribonucleoprotein – snRNP –snurp).Ø Ovi mnRNP čine jezgro čestica za isecanje.Ø RNK molekuli prepoznaju granice intron/egzon i učestvuju u reakcijamaisecanja.
Obrada RNK – isecanje intronaGUØ Tačku grananja prepoznaju BBP (branch pointbinding protein) i U2AF – pomoćni protein.Ø U2 mnRNP zamenjuje BBP i U2AF i baze mu sesparuju sa bazama konsenzus sekvence tačkegranjanja.Ø Baze U1 mnRNP se sparuju sa 5’ isecajućim mestom.Ø U4/U6 i U5 ulaze u česticu za isecanje.Ø U4 i U6 su međusobno čvrsto vezane preko parovabaza.Ø Dolazi do preuređenja RNK-RNK vezaAG
Obrada RNK – isecanje intronaØ U reakcijama transesterifikacije (transfosforilacije) broj fosfatnih veza ostaje isti,pa bi se moglo očekivati da se ove rekcije odvijaju bez hidrolize nukleozid trifosfata.Ø Međutim energija je potrebna za postepeno postavljanje i preuređenje komponenti čestica zaisecanje introna.- Neki od proteina koji ulaze u njihov sastav su helikaze koje koriste ATP za cepanje RNKRNK interakcija da bi omogućile stvaranje novih.- Hidrolizu ATP-a ne zahtevaju samoð Vezivanje BBP za tačku grananja ið Vezivanje U1 za 5’ isecajuće mesto.- Posle isecanja mnRNP ostaju vezani za omču introna, i rasformiranje ove strukturezahteva dodatna RNK-RNK preuređenja koja zahtevaju hidrolizu ATP-a, da bi mnRNK mogleda budu ponovo upotrebljene.Greške koje bi često nastajale da se selekcija mesta isecanjavrši na RNK bez proteina
Obrada RNK – isecanje intronaØIako se markiranje egzona i introna, i formiranje čestica za isecanje vršiu toku elongacije isecanje se vrši kasnije.ØOvo odlaganje znači da:ðse introni ne moraju isecati onim redom kojim su se pojavili u RNKlancuðIako se čestice za isecanje pojavljuju ko-transkripciono reakcijeisecanja se uglavnom odigravaju post-transkripciono, posle sintezekompletnog hnRNK
Obrada RNK – isecanje intronaØ Prve ćelije su upotrebljavale RNK molekule kaoglavne katalizatore i nosioce genetskihinformacija.Ø Dokaz za ovo shvatanje je postojanje samoisecajućih introna u nekih organizama(nuklearni rRNK geni trepljara Tetrahymena,nekoliko gena bakteriofaga T4) i u nekimgenima mitohondrija i hloroplasta.Ø Samo-isecanje introna se odigrava u odsustvuproteina i bilo kojih drugih RNK molekula.
Obrada RNK – isecanje intronaØ Dve grupe sekvenci samo-isecajućih introna:ð Grupa I sekvenci intronaIsecanje započinje vezivanjem Gnukleotida za sekvencu introna. G se takoaktivira i napada prvu fosfodiestarskuvezu (na 5’ mestu)ð Grupa II sekvenci intronaPosebno aktivan A u intronu je grupakoja napada i formira se intermedijeru vidu omče.Za obe grupe ključna je sekvenca introna.Smatra se da je isecanje hnRNK evoluiralo izklase II.
Transport iRNK iz jedraØ Transport zrele iRNK iz jedra u citoplazmu je visoko selektivan i zavisi odpravilne obrade hnRNK.Ø Kompleks nuklearnih pora prepoznaje i transportuje samo kompletnu iRNK.
rRNKLjudska ćelija sadrži oko 200 rRNK genskih kopija po haploidnom genomu, rasutih umalim grupama na pet različitih hromozoma.18S28S5.8S5S Ribozomalne RNK čine 80% RNK u ćeliji. 28S, 18S i 5.8S rRNK nastaju iz 45S prekursora rRNK koji se sintetiše u nukleolusu.Poreklo od istog gena obezbeđuje stvaranje sve tri rRNK u podjednakim količinama.5S rRNK nastaje u nukleoplazmi iz posebnog prekursora, koji se prepisuje sadrugog gena.
Sinteza i obrada rRNKØ rRNK je najzastupljenija RNK u ćeliji – oko 80% ukupne RNK u ćeliji koja se brzo deli.Ø Sintetiše ih RNK polimeraza I (28S, 18S, 5.8S) i RNK polimeraza III (5S).Ø Slična je RNK polimerazi II ali nema C-terminalni “rep“ što objašnjava zašto rRNKnema ni 5’ “kapu“ ni poli-A “ rep “.Ø Rastuća ćelija sisara mora da sintetiše oko 10 miliona kopija svakog tipa rRNK u svakojćelijskoj generaciji.Ø Humana ćelija sadrži oko 200 rRNK gena u haploidnom genomu raspoređenih na 5hromozoma.5’45S prekursor rRNK3’
Sinteza i obrada rRNKØ Prekursor rRNK je dug oko 13.000 nukleotida.Ø Veliki broj modifikacija se odigrava pre njegovogisecanja:- Oko 100 reakcija metilacije na 2’ OH šećera- Oko 100 izomerizacija uridina u pseudouridinØ Svaka modifikacija se odvija na određenoj pozicijirRNK.Ø Pozicije su određene pomoću specifičnih RNK kojese nazivaju male nukleolarne RNK(small nucleolar RNA - snoRNAs)- Bazno se sparuju sa prekursornom rRNK i- Postavljaju RNK-modifikujuće enzime uodgovarajućem položaju- uzrokuju konformacione promene prekursornerRNK i dovode do cepanja prekursora u zrelerRNK.-Mnoge snoRNA kodiraju introni drugih gena,posebno onih koji kodiraju ribozomalne proteine.
rRNK strukturasekundarnaPretpostavljena sekundarna struktura za 5S rRNK.
Sinteza i obrada rRNKNukleolusØ Nije ograničen membranomØ Veliki agregat makromolekulað rRNK genið Prekursori rRNKð Zrela rRNKð Enzimi obrade rRNKð snoRNPð Proteinske subjedinice ribozomað Delimično sklopljeni ribozomiØ Osim ribozoma u nukleolusu se stvaraju idruge RNK i RNK-protein kompleksið U6 mnRNPð telomerazað čestica za prepoznavanje signalað tRNK
tRNK - sekundarna strukturaMesto vezivanjaamino kiselineDHU petljaTψC petljaSadrži dihidrouracilMesto vezivanjaamino acil-tRNK transferazeUčestvuje u vezivanjuza ribozomAntikodonska petljaPrepoznaje kodon na i RNK
Sinteza i obrada tRNKØ Transportna RNK je mali molekul veličine 70-90 baza.Ø Nastaje isecanjem iz velikih primarnih prekursora,koji sadrže sekvence za 2 do 7 molekula tRNK.Ø Isecanje funkcionalnih delova tRNK katališu enzimiribonukleaza P i ribonukleaza D.- Ribonukleaza P endonukleaznim cepanjem odvaja 5’ kraj,a 3’ kraj se odvaja egzonukleaznim cepanjem.Ø Posle odvajanja tRNK, na 3’ kraju počinje sintezaCCA sekvence, katalisana tRNK nukleotidiltransferazom.Ø Dalja obrada molekula tRNK obuhvata modifikovanje baza.- Postoji preko 60 različitih modifikacija, od kojih jenajčešća metilacija. Većina baza tRNK su metilovane.Ø Neke tRNK sadrže male introne (14-50 nukleotida) kojese uklanjaju isecanjem i ponovnim povezivanjem RNK
Neouobičajeni NT prisutni u molekulu tRNKOvi nukleotidi nastaju kovalentnom modifikacijom normalnihnukleotida po njihovom ugrađivanju u lanac RNK.U celoj tRNK, modifikovano je oko 10% nukleotida
Mitohondrijalni genom
Mitohondrijalni genomTeški lanacLaki lanacOko 17000bp, 37 genaSadrži malo nekodirajućih sekvenciSopstvena replikacija i traskripcija
završetak sinteze molekula RNK. Ø Ispred mesta terminacije postoji sekvenca bogata citozinom i guaninom - palindrom sekvenca, koja sadrži međusobno komplementarne regione, dolazi do savijanja RNK molekula i formiranja petlje u obliku ukosnice. Ø RNK polimeraza se zaustavlja i ponovo se formira DNK heliks Ukosnica -terminacioni signal iRNK