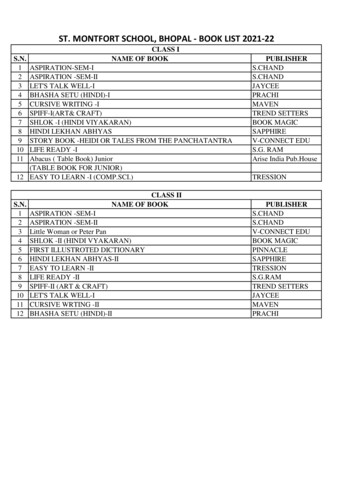
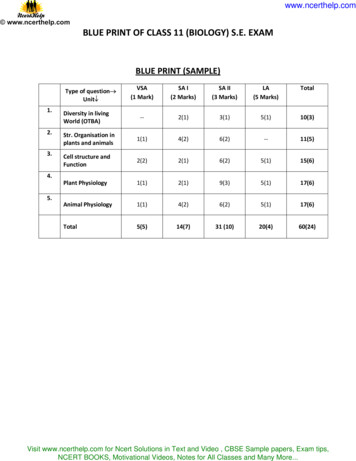
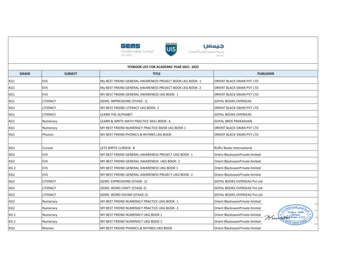
Transcription
CHAPTER9Heredity andEvolutionWe have seen that reproductive processes give rise to new individualsthat are similar, but subtly different. We have discussed how someamount of variation is produced even during asexual reproduction. Andthe number of successful variations are maximised by the process ofsexual reproduction. If we observe a field of sugarcane we find very littlevariations among the individual plants. But in a number of animalsincluding human beings, which reproduce sexually, quite distinctvariations are visible among different individuals. In this chapter, weshall be studying the mechanism by which variations are created andinherited. The long-term consequences of the accumulation of variationsare also an interesting point to be considered. We shall be studying thisunder evolution.9 . 1 A CCUMUL ATION OF VARIATIONVARIAARIATIONDURING REPRODUCTIONFigure 9.1Creation of diversity over succeedinggenerations. The original organism at the topwill give rise to, say, two individuals, similarin body design, but with subtle differences.Each of them, in turn, will give rise to twoindividuals in the next generation. Each ofthe four individuals in the bottom row will bedifferent from each other. While some of thesedifferences will be unique, others will beinherited from their respective parents, whowere different from each other.142Inheritance from the previous generation provides botha common basic body design, and subtle changes in it,for the next generation. Now think about what wouldhappen when this new generation, in its turn,reproduces. The second generation will have differencesthat they inherit from the first generation, as well asnewly created differences (Fig. 9.1).Figure 9.1 would represent the situation if a singleindividual reproduces, as happens in asexualreproduction. If one bacterium divides, and then theresultant two bacteria divide again, the four individualbacteria generated would be very similar. There wouldbe only very minor differences between them, generateddue to small inaccuracies in DNA copying. However, ifsexual reproduction is involved, even greater diversitywill be generated, as we will see when we discuss therules of inheritance.Do all these variations in a species have equalchances of surviving in the environment in which theyfind themselves? Obviously not. Depending on thenature of variations, different individuals would haveScience
different kinds of advantages. Bacteria that can withstand heat will survivebetter in a heat wave, as we have discussed earlier. Selection of variantsby environmental factors forms the basis for evolutionary processes, aswe will discuss in later sections.QUESTIONS1.If a trait A exists in 10% of a population of an asexually reproducingspecies and a trait B exists in 60% of the same population, which traitis likely to have arisen earlier?2.How does the creation of variations in a species promote survival?9.2 HEREDITYThe most obvious outcome of the reproductive process still remains thegeneration of individuals of similar design. The rules of heredity determinethe process by which traits and characteristics are reliably inherited. Letus take a closer look at these rules.9.2.1 Inherited TraitsWhat exactly do we mean by similarities and differences? We know thata child bears all the basic features of a human being. However, it doesnot look exactly like its parents, and human populations show a greatdeal of variation.Activity 9.1 Observe the ears of all the students in the class. Prepare a list ofstudents having free or attached earlobes and calculate thepercentage of students having each (Fig. 9.2). Find out about theearlobes of the parents of each student in the class. Correlate theearlobe type of each student with that of their parents. Based onthis evidence, suggest a possible rule for the inheritance of earlobetypes.9.2.2 Rules for the Inheritance of Traits –Mendel’s ContributionsThe rules for inheritance of such traits in human beings are related tothe fact that both the father and the mother contribute practically equalamounts of genetic material to the child. This means that each trait canbe influenced by both paternal and maternal DNA. Thus, for each traitthere will be two versions in each child. What will, then, the trait seen inthe child be? Mendel (see box) worked out the main rules of suchinheritance, and it is interesting to look at some of his experiments frommore than a century ago.Heredity and Evolution(a)(b)Figure 9.2(a) Free and (b) attachedearlobes. The lowest partof the ear, called theearlobe, is closely attachedto the side of the head insome of us, and notin others. Free andattached earlobes are twovariants found in humanpopulations.143
Gregor Johann Mendel (1822–1884)Mendel was educated in a monastery and went on to study science andmathematics at the University of Vienna. Failure in the examinations for ateaching certificate did not suppress his zeal for scientific quest. He wentback to his monastery and started growing peas. Many others had studiedthe inheritance of traits in peas and other organisms earlier, but Mendelblended his knowledge of science and mathematics and was the first oneto keep count of individuals exhibiting a particular trait in each generation.This helped him to arrive at the laws of inheritance that we have discussedin the main text.Mendel used a number of contrasting visible characters of gardenpeas – round/wrinkled seeds, tall/short plants, white/violet flowers andso on. He took pea plants with different characteristics – a tall plant anda short plant, produced progeny from them, and calculated thepercentages of tall or short progeny.In the first place, there were no halfway characteristics in this firstgeneration, or F1 progeny – no ‘medium-height’ plants. All plants weretall. This meant that only one of the parental traitswas seen, not some mixture of the two. So the nextquestion was, were the tall plants in the F1generation exactly the same as the tall plants of theparent generation? Mendelian experiments test thisby getting both the parental plants and these F1 tallplants to reproduce by self-pollination. The progenyof the parental plants are, of course, all tall. However,the second-generation, or F2, progeny of the F1 tallplants are not all tall. Instead, one quarter of themare short. This indicates that both the tallness andshortness traits were inherited in the F1 plants, butonly the tallness trait was expressed. Thus, twocopies of the trait are inherited in each sexuallyreproducing organism. These two may be identical,or may be different, depending on the parentage. Apattern of inheritance can be worked out with thisassumption, as shown in Fig. 9.3.Figure 9.3Inheritance of traitsover two generationsActivity 9.2 In Fig. 9.3, what experiment would we do to confirm that the F2generation did in fact have a 1:2:1 ratio of TT, Tt and tt traitcombinations?In this explanation, both TT and Tt are tall plants, while only tt is ashort plant. In other words, a single copy of ‘T’ is enough to make theplant tall, while both copies have to be ‘t’ for the plant to be short. Traitslike ‘T’ are called dominant traits, while those that behave like ‘t’ arecalled recessive traits. Work out which trait would be considereddominant and which one recessive in Fig. 9.4.144Science
What happens when pea plants showing two differentcharacteristics, rather than just one, are bred with each other?What do the progeny of a tall plant with round seeds and a shortplant with wrinkled-seeds look like? They are all tall and haveround seeds. Tallness and round seeds are thus dominant traits.But what happens when these F1 progeny are used to generateF2 progeny by self-pollination? A Mendelian experiment will findthat some F2 progeny are tall plants with round seeds, and somewere short plants with wrinkled seeds. However, there wouldalso be some F2 progeny that showed new mixtures. Some ofthem would be tall, but have wrinkled seeds, while others wouldbe short, but have round seeds. Thus, the tall/short trait andthe round seed/wrinkled seed trait are independently inherited.Another example is shown in Fig. 9.5.Figure 9.49.2.3 How do these Traits get Expressed?How does the mechanism of heredity work? Cellular DNA isthe information source for making proteins in the cell. Asection of DNA that provides information for one protein iscalled the gene for that protein. How do proteins control thecharacteristics that we are discussing here? Let us take theexample of tallness as a characteristic. We know that plantshave hormones that can trigger growth. Plant height canthus depend on the amount of a particular plant hormone.The amount of the plant hormone made will depend on theefficiency of the process for making it. Consider now anenzyme that is important for this process. If this enzymeworks efficiently, a lot of hormone will be made, and theplant will be tall. If the gene for that enzyme has an alterationthat makes the enzyme less efficient, the amount of hormonewill be less, and the plant will be short. Thus, genes controlcharacteristics, or traits.If the interpretations of Mendelian experiments we havebeen discussing are correct, then both parents must becontributing equally to the DNA of the progeny during sexualreproduction. We have disscussed this issue in the previousChapter. If both parents can help determine the trait in theprogeny, both parents must be contributing a copy of the samegene. This means that each pea plant must have two sets of allgenes, one inherited from each parent. For this mechanism towork, each germ cell must have only one gene set.How do germ-cells make a single set of genes from thenormal two copies that all other cells in the body have? If progeny plantsinherited a single whole gene set from each parent, then the experimentin Fig. 9.5 cannot work. This is because the two characteristics ‘R’ and‘y’ would then be linked to each other and cannot be independentlyinherited. This is explained by the fact that each gene set is present, notHeredity and EvolutionFigure 9.5Independent inheritanceof two separate traits,shape and colour of seeds145
as a single long thread of DNA, but as separate independent pieces,each called a chromosome. Thus, each cell will have two copies of eachchromosome, one each from the male and female parents. Every germcell will take one chromosome from each pair and these may be of eithermaternal or paternal origin. When two germ cells combine, they willrestore the normal number of chromosomes in the progeny, ensuringthe stability of the DNA of the species. Such a mechanism of inheritanceexplains the results of the Mendel experiments, and is used by allsexually reproducing organisms. But asexually reproducing organismsalso follow similar rules of inheritance. Can we work out how theirinheritance might work?9.2.4 Sex DeterminationFigure 9.6Sex determination inhuman beings146We have discussed the idea that the two sexes participating in sexualreproduction must be somewhat different from each other for a numberof reasons. How is the sex of a newborn individualdetermined? Different species use very different strategiesfor this. Some rely entirely on environmental cues. Thus,in some animals, the temperature at which fertilised eggsare kept determines whether the animals developing inthe eggs will be male or female. In other animals, such assnails, individuals can change sex, indicating that sex isnot genetically determined. However, in human beings,the sex of the individual is largely genetically determined.In other words, the genes inherited from our parentsdecide whether we will be boys or girls. But so far, wehave assumed that similar gene sets are inherited fromboth parents. If that is the case, how can geneticinheritance determine sex?The explanation lies in the fact that all humanchromosomes are not paired. Most human chromosomeshave a maternal and a paternal copy, and we have 22such pairs. But one pair, called the sex chromosomes, isodd in not always being a perfect pair. Women have aperfect pair of sex chromosomes, both called X. But menhave a mismatched pair in which one is a normal-sized Xwhile the other is a short one called Y. So women are XX,while men are XY. Now, can we work out what theinheritance pattern of X and Y will be?As Fig. 9.6 shows, half the children will be boys andhalf will be girls. All children will inherit an X chromosomefrom their mother regardless of whether they are boys orgirls. Thus, the sex of the children will be determined bywhat they inherit from their father. A child who inheritsan X chromosome from her father will be a girl, and onewho inherits a Y chromosome from him will be a boy.Science
QUESTIONS1.How do Mendel’s experiments show that traits may be dominant orrecessive?2.How do Mendel’s experiments show that traits are inheritedindependently?3.A man with blood group A marries a woman with blood group O andtheir daughter has blood group O. Is this information enough to tell youwhich of the traits – blood group A or O – is dominant? Why or why not?4.How is the sex of the child determined in human beings?9.3 EVOLUTIONWe have noted that there is an inbuilt tendency to variation duringreproduction, both because of errors in DNA copying, and as a result ofsexual reproduction. Let us now look at some consequences of thistendency.9.3.1 An IllustrationConsider a group of twelve red beetles. They live, let us assume, in somebushes with green leaves. Their population will grow by sexualreproduction, and therefore, can generate variations. Let us imagine alsothat crows eat these beetles. The more beetles the crows eat, the fewerbeetles are available to reproduce. Now, let us think about some differentsituations (Fig. 9.7) that can develop in this beetle population.Figure 9.7 Variations in a population – inherited and otherwiseIn the first situation, a colour variation arises during reproduction,so that there is one beetle that is green in colour instead of red. Thisbeetle, moreover, can pass the colour on to its progeny, so that all itsHeredity and Evolution147
progeny beetles are green. Crows cannot see green-coloured beetles onthe green leaves of the bushes, and therefore cannot eat them. Whathappens then? The progeny of green beetles is not eaten, while theprogeny of red beetles continues to be eaten. As a result, there are moreand more green beetles than red ones in the beetle population.In a second situation, again, a colour variation arises duringreproduction, but now it results in a beetle that is blue in colour insteadof red. This beetle can also pass the colour on to its progeny, so that allits progeny beetles are blue. Crows can see blue-coloured beetles in thegreen leaves of the bushes as well as they can see red ones, and thereforecan eat them. What happens initially? In the population, as it expands,there are a few blue beetles, but most are red. But at this point, anelephant comes by, and stamps on the bushes where the beetles live.This kills most of the beetles. By chance, the few beetles that have survivedare mostly blue. The beetle population slowly expands again, but now,the beetles in the population are mostly blue.It is obvious that in both situations, what started out as a rare variationcame to be a common characteristic in the population. In other words,the frequency of an inherited trait changed over generations. Since genescontrol traits, we can say that the frequency of certain genes in apopulation changed over generations. This is the essence of the idea ofevolution.But there are interesting differences, too, in the two situations. In thefirst case, the variation became common because it gave a survivaladvantage. In other words, it was naturally selected. We can see that thenatural selection is exerted by the crows. The more crows there are, themore red beetles would be eaten, and the more the proportion of greenbeetles in the population would be. Thus, natural selection is directingevolution in the beetle population. It results in adaptations in the beetlepopulation to fit their environment better.In the second situation, the colour change gave no survival advantage.Instead, it was simply a matter of accidental survival of beetles of onecolour that changed the common characteristic of the resultantpopulation. The elephant would not have caused such major havoc inthe beetle population if the beetle population had been very large. So,accidents in small populations can change the frequency of some genesin a population, even if they give no survival advantage. This is the notionof genetic drift, which provides diversity without any adaptations.Now consider a third situation. In this, as the beetle population beginsto expand, the bushes start suffering from a plant disease. The amountof leaf material for the beetles is reduced. The beetles are poorly nourishedas a result. The average weight of adult beetles decreases from what itused to be when leaves were plentiful, but there is no genetic changeoccurring. After a few years and a few beetle generations of such scarcity,the plant disease is eliminated. There is a lot of leaf food. At this time,what would we expect the weight of the beetles to be?148Science
9.3.2 Acquired and Inherited TraitsWe discussed the idea that the germ cells of sexually reproducingpopulations are made in specialised reproductive tissue. If the weight ofthe beetle is reduced because of starvation, that will not change the DNAof the germ cells. Therefore, low weight is not a trait that can be inheritedby the progeny of a starving beetle. Therefore, even if some generationsof beetles are low in weight because of starvation, that is not an exampleof evolution, since the change is not inherited over generations. Changein non-reproductive tissues cannot be passed on to the DNA of the germcells. Therefore the experiences of an individual during its lifetime cannotbe passed on to its progeny, and cannot direct evolution.Consider another example of how an individual cannot pass on toits progeny the experiences of its lifetime. If we breed a group of mice, alltheir progeny will have tails, as expected. Now, if the tails of these miceare removed by surgery in each generation, do these tailless mice havetailless progeny? The answer is no, and it makes sense because removalof the tail cannot change the genes of the germ cells of the mice.Charles Robert Darwin (1809–1882)Charles Darwin set out on a voyage when he was 22 years old. Thefive-year voyage took him to South America and the islands off its coast.The studies that he conducted during this voyage were to change foreverthe way we look at the variety of life on earth. Interestingly, after he gotback to England, he never left its shores again. He stayed at home andconducted various experiments that led him to formulate his hypothesisthat evolution took place due to natural selection. He did not know themechanism whereby variations arose in the species. He would havebeen enlightened by Mendel’s experiments, but these two gentlemendid not know of each other or their work!We often associate Darwin solely with the theory of evolution. But he was an accomplishednaturalist, and one of the studies he conducted was to do with the role of earthworms insoil fertility.This is the reason why the ideas of heredity and genetics that wehave discussed earlier are so essential for understanding evolution. EvenCharles Darwin, who came up with the idea of evolution of species bynatural selection in the nineteenth century, could not work out themechanism. It is ironic that he could have done so if he had seen thesignificance of the experiments his Austrian contemporary, GregorMendel, was doing. But then, Mendel too did not notice Darwin’s workas relevant to his!Origin of life on earthDarwin’s theory of evolution tells us how life evolved from simple to more complex formsand Mendel’s experiments give us the mechanism for the inheritance of traits from onegeneration to the next. But neither tells us anything about how life began on earth in thefirst place.Heredity and Evolution149
Do You Know?J.B.S. Haldane, a British scientist (who became a citizen of India later), suggested in1929 that life must have developed from the simple inorganic molecules which werepresent on earth soon after it was formed. He speculated that the conditions on earthat that time, which were far from the conditions we see today, could have given rise tomore complex organic molecules that were necessary for life. The first primitiveorganisms would arise from further chemical synthesis.How did these organic molecules arise? An answer was suggested by the experimentconducted by Stanley L. Miller and Harold C. Urey in 1953. They assembled anatmosphere similar to that thought to exist on early earth (this had molecules likeammonia, methane and hydrogen sulphide, but no oxygen) over water. This wasmaintained at a temperature just below 100 C and sparks were passed through themixture of gases to simulate lightning. At the end of a week, 15% of the carbon (frommethane) had been converted to simple compounds of carbon including amino acidswhich make up protein molecules. So, can life arise afresh on earth even now?QUESTIONS1.What are the different ways in which individuals with a particulartrait may increase in a population?2.Why are traits acquired during the life-time of an individual notinherited?3.Why are the small numbers of surviving tigers a cause of worry fromthe point of view of genetics?9.4 SPECIATIONSPECIATIONWhat we have seen so far is micro-evolution. That means that the changesare small, even though they are significant. Also, they simply change thecommon characteristics of a particular species. But this does not properlyexplain how new species come into existence. That can be said to havehappened only if this group of beetles we are thinking about, splits intotwo populations that cannot reproduce with each other. When thishappens, they can be called two independent species. So, can we extendthe reasoning we have used above to explain such speciation?Consider what would happen if the bushes the beetles feed on arespread widely over a mountain range. The beetle population becomesvery large as a result. But individual beetles feed mostly on a few nearbybushes throughout their lifetime. They do not travel far. So, in this hugepopulation of beetles, there will be sub-populations in neighbourhoods.Since male and female beetles have to meet for reproduction to happen,most reproduction will be within these sub-populations. Of course, anoccasional adventurous beetle might go from one site to another. Or abeetle is picked up by a crow from one site and dropped in the other sitewithout being eaten. In either case, the migrant beetle will reproducewith the local population. This will result in the genes of the migrantbeetle entering a new population. This kind of gene flow is bound to150Science
happen between populations that are partly, but not completelyseparated. If, however, between two such sub-populations a large rivercomes into existence, the two populations will be further isolated. Thelevels of gene flow between them will decrease even further.Over generations, genetic drift will accumulate different changes ineach sub-population. Also, natural selection may also operate differentlyin these different geographic locations. Thus, for example, in the territoryof one sub-population, crows are eliminated by eagles. But this doesnot happen for the other sub-population, where crow numbers are veryhigh. As a result, the green variation will not be selected at the first site,while it will be strongly selected at the second.Together, the processes of genetic drift and natural selection will resultin these two isolated sub-populations of beetles becoming more andmore different from each other. Eventually, members of these two groupswill be incapable of reproducing with each other even if they happen tomeet.There can be a number of ways by which this can happen. If theDNA changes are severe enough, such as a change in the number ofchromosomes, eventually the germ cells of the two groups cannot fusewith each other. Or a new variation emerges in which green females willnot mate with red males, but only with green males. This allows verystrong natural selection for greenness. Now, if such a green female beetlemeets a red male from the other group, her behaviour will ensure thatthere is no reproduction between them. Effectively, new species of beetlesare being generated.QUESTIONS1.What factors could lead to the rise of a new species?2.Will geographical isolation be a major factor in the speciation of a selfpollinating plant species? Why or why not?3.Will geographical isolation be a major factor in the speciation of anorganism that reproduces asexually? Why or why not?9.5 EVOLUTION AND CL ASSIFIC ATIONEVOLUTIONBased on these principles, we can work out the evolutionary relationshipsof the species we see around us. It is a sort of going backwards in time.We can do this by identifying hierarchies of characteristics betweenspecies. In order to understand this process, let us think back to ourdiscussion on the classification of organisms in Class IX.Similarities among organisms will allow us to group them and thenstudy the groups. For this, which characteristics decide morefundamental differences among organisms, and which ones decide lessbasic differences? What is meant by ‘characteristics’, anyway?Characteristics are details of appearance or behaviour; in other words, aparticular form or a particular function. That we have four limbs is thus acharacteristic. That plants can do photosynthesis is also a characteristic.Heredity and Evolution151
Some basic characteristics will be shared by most organisms. Thecell is the basic unit of life in all organisms. The characteristics in thenext level of classification would be shared by most, but not all organisms.A basic characteristic of cell design that differs among different organismsis whether the cell has a nucleus. Bacterial cells do not, while the cells ofmost other organisms do. Among organisms with nucleated cells, whichones are unicellular and which ones multi-cellular? That property marksa very basic difference in body design, because of specialisation of celltypes and tissues. Among multi-cellular organisms, whether they canundertake photosynthesis or not will provide the next level ofclassification. Among the multi-cellular organisms that cannot dophotosynthesis, whether the skeleton is inside the body or around thebody will mark another fundamental design difference. We can see that,even in these few questions that we have asked, a hierarchy is developingthat allows us to make classification groups.The more characteristics two species will have in common, the moreclosely they are related. And the more closely they are related, the morerecently they will have had a common ancestor. An example will help. Abrother and a sister are closely related. They have common ancestors inthe first generation before them, namely, their parents. A girl and herfirst cousin are also related, but less than the girl and her brother. Thisis because cousins have common ancestors, their grandparents, in thesecond generation before them, not in the first one. We can now appreciatethat classification of species is in fact a reflection of their evolutionaryrelationship.We can thus build up small groups of species with recent commonancestors, then super-groups of these groups with more distant commonancestors, and so on. In theory, we can keep going backwards like thisuntil we come to the notion of a single species at the very beginning ofevolutionary time. If that is the case, then at some point in the history ofthe earth, non-living material must have given rise to life. There are manytheories about how this might have happened. It would be interesting tocome up with theories of our own!9.5.1 Tracing Evolutionary RelationshipsFigure 9.8Homologous organs152When we try to follow evolutionary relationships, how do we identifycharacteristics as common? These characteristics in different organismswould be similar because they are inherited from a common ancestor.As an example, consider the fact that mammals have four limbs, as dobirds, reptiles and amphibians (Fig. 9.8). The basic structure of the limbsis similar though it has been modified to perform different functions invarious vertebrates. Such a homologous characteristic helps to identifyan evolutionary relationship between apparently different species.However, all similarities simply in organ shape are not necessarilybecause of common ancestry. What would we think about the wings ofScience
birds and bats, for example (Fig. 9.9)? Birdsand bats have wings, but squirrels andlizards do not. So are birds and bats moreclosely related to each other than to squirrelsor lizards?Before we jump to this conclusion, let uslook at the wings of birds and bats moreFigure 9.9closely. When we do that, we find that theAnalogous organs – The wing of a bat and thewings of bats are skin folds stretched mainlywing of a birdbetween elongated fingers. But the wings ofbirds are a feathery covering all along the arm. The designs of the twowings, their structure and components, are thus very different. Theylook similar because they have a common use for flying, but their originsare not common. This makes them analogous characteristics, ratherthan homologous characteristics. It would now be interesting to thinkabout whether bird arms and bat arms should be considered homologousor analogous!9.5.2 FossilsSuch studies of organ structure can be done not only on current species,but also on species that are no longer alive. How do we know that theseextinct species ever existed? We know this from finding fossils (Fig. 9.10).What are fossils? Usually, when organisms die, their bodies willdecompose and be lost. But every once in a while, the body or at leastsome
the child be? Mendel (see box) worked out the main rules of such inheritance, and it is interesting to look at some of his experiments from more than a century ago. Figure 9.2 (a) Free and (b) attached earlobes. The lowest part of the ear, called the earlobe, is closely attached to the side of the head in